|
Cognitive
Systems 1/4, S. 305-320
1986
On
Concepts of "Cognition" in Biology
Abstract
Besides
an introductory chapter on "cognitive" processes in the
biomolecular domain (part A), this essay presents some reflections
on vertebrate behaviour from a neurocybernetic point of view. First,
some comments are given on the problem of modeling organism-environment
interactions (part B).
It is shown how environmental features enter the nervous system
by altering a behaviour-immanent sensorimotor circuit.
In
following a coarse phylogenetic line - with only consideration of
three major stages (fish, tetrapodes, and primates) - it is argued
that extensions of the sensorimotor circuits are parallelled by
increasing complexity of locomotor behaviour, part C.
The
last part, D, deals with permanent bipedality of hominids and points
to two consequences of the fact that arms and hands lost their locomotor
function: (i) a compensatory drive towards object manipulations,
and (ii) an additional quality ("Eigenquality") in experiencing
own manipulations under visual selfcontrol.
The
paper is intended to support a provoking hypothesis: Could the origin
of aspects of selfconsciousness have been just (due to) an extra
control loop of arm and hand movements?
"Even
the highest evolved nervous system
can never be wiser than its Afferenzen
(afferent informations) permit"
(E. von Holst, 1950)
Part
A. On behavior of biomolecules
If we believe in evolution, we accept that live originated in an
aqueous environment. Whether we take a cell. as the minimum condition
which shows all categories of life phenomena, or even a precellular,
molecular phase, makes no difference: if there was cognition it
was without psychology.
Biologists live largely within a man-made environment; the main
problem of dealing with cognition at the infrahuman level will be
to get rid of anthropomorphism. Did Konrad Lorenz do a big step
forward (and backwards along evolutionary lines) by saying: "Life
itself is a cognitive process" ? (1973). Similarly, Humberto
Maturana stated: "Living systems are cognitive systems, and
to live is to know." (private communication).
Even if we try to look for the physical forces involved in metabolism,
no answer to the question of the appropriateness of the term "cognition"
is given. Two sorts of forces help to bring about such "cognitive"
processes:
1. Thermodynamic motion mixes the interactions in a chaotic manner
until complementary structures "catch each other".
2. The "catching" is the sum of electrostatic forces exerted
between pattern-matching surfaces. (Chapeville e.a. 1980)
Cognition
at the molecular level can thus be seen as biophysicalchemical interactions,
selective by pattern-matching between "spatially complementary"
structures (note 1). Water, an ever-changing medium providing a
lot of variety of interaction partners in rapid succession, is the
necessary precondition.
Now let us embark on the stream of evolution. After development
of protective double layers, the cell shielded off hydrolytic influence
from the aqueous environment. Catalytic interactions now worked
within a spatially autonomous, but energetically and informationally
open system. Food recognition and communication (sexuality !) were
still mediated by molecular interaction with receptors "waiting"
an the cell surface. Our chemical senses (taste and smell), hormone
effects, and immunological "recognition", all work on
the direct-contact-complementarity principle.
Contact problems had to be solved, too, when cell colonies appeared
and started a new, most sucessful level of life phenomena, that
of multicellular organisms, the metazoans.
Size and volume of higher metazoans prevented external thermodynamic
motion forces to push them around. Metazoans had either to stick
to the ground (plants, and sessile animals), float more or less
freely (e.g. some cnidarians), or invent locomotion.
Part
B. The basic sensorimotor system
"Muscle
moves the world" (Ragnar Granit)
"Behaviour is regular, but there are no regulators" (J.
Gibson)
B1.
Beyond a stimulus-response model
Organismic
movement must not be random. Orientation must bring about the connection
between organism and its needs (food, shelter, partners). This is
where sense organs (note 2) are necessary, with connections to the
nervous system, making it a central nervous system (CNS).
Let us describe that system in more detail. It consists of two sorts
of elements:
|
Material
elements:
- a
brain, or central nervous system, CNS,
- afferent
connections, leading from sense organs to the CNS,
- efferent
connections leading from the CNS to muscles and glands,
- association
fibres,
- connections
within the brain.
|
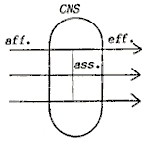
Fig.1. Material elements |
|
Dynamic elements:
- efferent
motor patterns, which are excitation patterns for muscle innervations
traveling from the CNS to the "effector" muscles,
and
- afferent
sensory patterns, excitation patterns from the receptors (on the
surface or within the organism) to the CNS
Apparently
the motor apparatus is, at least partially, triggered from outside.
The role of a CNS seems to be the coordination
of stimuli from the environment
(note 3) with movements. Behavior was never independent
of sensory input. Such a description shows
how spontaneous movements can be explained as reactions,
see Fig. 2.
|
Fig.
2.
Stimuli and reactions, interactions
with the environment
(Figure after Ingenieros 1922)
|
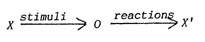
X =
environment, 0= organism
|
The
stimulus-response model of classical behaviorism can be easily recognized,
which gave rise to abundant literature. An experimenter can administer
artificial stimuli without bounds, and so manipulate resulting behaviour.
We shall now introduce a modified model, in order to avoid some serious
disadvantages. Part of the modification consists in adding some trivial
assumptions. Let us consider a hypothetical palaezoic fish as a starting
point, to make the discussion more concrete. (We avoid speculations
on the origin of vertebrates)
The basic task of the CNS is the coordination of locomotion with sensory
input. The critical, but trivial assumption is to recognize that an
organism by locomotion changes its environment
Ashby 1952: p.37: "...muscles affect the receptors (by effects
transmitted through the environment)..." "...most physiological
experiments are deliberately arranged to avoid this feedback".
One could say that by locomotion the organism stimulates its own sense
organs. Or, alternatively: sensory input is not independent of behavior
like locomotion. |
| Fig.
3. The organism (0) acting on the environment (X),
and sensing it. |
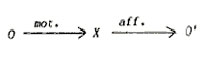 |
B2.
Beyond a "straight-line" model
In vertebrates important sense organs are situated at the front of
the organism. Locomotion is direction-selective, and constrained.
This may have been the reason for the development of the CNS in the
front position. The diagram of information channels may then be drawn
in terms of loops: |
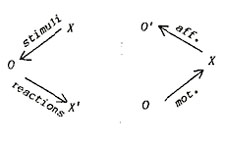 |
X,
X´= environment, experimenter
O, O´= organism, CNS
Fig. 4.
Two kinds of interactions with the environment.
|
| If we
join the behaviorist loop (left) and the self-stimulative loop (right),
we obtain a closed loop, which can serve as the basis of an orientational
system |
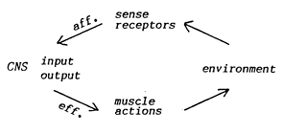 |
Fig.
5. An orientational system. |
Several
disadvantages make a drawing of a circuit a bad model for environment-related
behaviour:
a. The system is not a closed material circuit, it consists of various
dynamic processes chained together. It runs as follows: motor neuronal
discharge - transmission along motor pathways - contraction of muscles
- displacement of the organism in the environment - perception of
a changing environment - transmission along sensory pathways - feeding
into the CNS, convergence (in yet unidentified ways) an the motor
areas - activation of motor programs.
b. Environment change is subjective experience, an external observer
can only describe an organism in locomotion, he can only imagine an
"environment-shift-effect" for the observed organism.
Experiences attributed to another subject without verifiability for
the observer, that is where psychology begins.
c. Motor pathways leave the brain in nicely ordered, parallel fibres,
sensory input is dispersed, coming from many directions. Often several
sensory responses (vestibular (note 4), visual, tactile, chemical)
are fed back simultaneously, Fig. 6. |
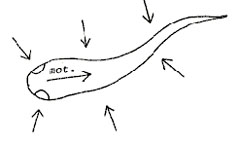 |
Fig.
6. Multiple sensory responses.
|
| d. Regarding
the multiple feedbacks from motor activity, the system can be viewed
as an amplifier, regarding the vast input and the comparably "thin"
output, it can be viewed as a filter: |
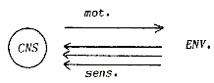 |
Fig.
7. Multiple feedbacks.
|
|
e.
The explanation of adaptation is a big problem. Apparently, the
CNS can use its afferent information to react in an appropriate,
oriented manner to environmental features. We call the behavior
of animals adaptive, often goal-directed. If so, the vast amount
of sensory input must be filtered, selected, reduced, and finally
be used as triggers and regulators for the motor apparatus.
According to J. Gray (1950), motor control in vertebrates seems
to have proceeded from an endogeneous, preprogrammed, largely rhythmic
type, as described for fish by E. von Holst (1969,1970), to an increasingly
modifiable type of motor control dependent an complex sensory processing.
From the brains of higher vertebrates there is anatomical evidence:
the structures doing the information-processing of sensory input
appear to have "overwhelmed" the motor apparatus.
The
organism-environment interaction as a circular interaction has been
introduced before by Victor von Weizsäcker and Jacob von Uexkuell.
The first wrote a monograph "Der Gestaltkreis" undertitled
"A theory about the unity of perception and movements".
His figure reads as follows:
|
0=
Organism
U= Umwelt = environment |
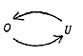 |
|
Fig.
8. Complementarity of organism and environment.
|
|
Being
a neurologist, von Weizsäcker noticed complementarity in movement-perception
acts and states a mutual exclusion-principle insofar as perception
can be replaced by spontaneous movement (and vice versa). He thus
drew attention to the inseparability of movement and perception:
"sensomobility".
More
elaborate is the Funktionskreis by von Uexkuell, a famous biologist.
His figure:
|
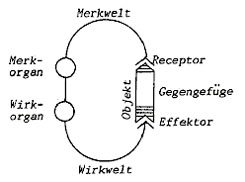 |
Merkwelt= "world of perception"
Wirkwelt= "world of action"
Gegengefüge = environment as a structured whole |
| Fig.
9. Funktionskreis by von Uexkuell (1930) |
His
basic metaphor for the circuit is "Zange" (a pair of pliers),
by the Funktionskreis the subject grasps the object.
The following two paragraphs are slightly corrected for clearer understanding
(edited and added 2015)
B3. Towards a dynamic model
We use a tree model to show the organization of our (human) NS but to arrive at it we have to turn the basic sensori-motor (or kin-esthetic) loop/circuit on its head.
Showing the circuit “upside down” is defamiliarizing but at the same time it can help by the tree analogy – a fountain would do even better, by adding the temporal domain:
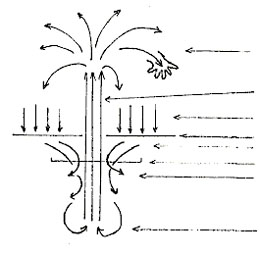 |
Peripheral motor nerves
tractus corticospinalis
stimuli
sensory surfaces: skin, eye
peripheral sensory nerves
medulla spinalis
sensory-motor-counter flow
"motor control"
cortex: sensory-motor association areas
|
Fig.10.
The stem is the main motor output of the CNS/brain, the corticospinal tract leaving the head through the occipital foramen (in reality “down”wards) into the vertebrate column. In each intervertebral joint motor nerves sprout to the side innervating the arms and legs – thereby spreading out to the physical world, towards objects etc.
Touching means the objects push back, and this ”echo” of the motor pulses “comes back” through the skin - now called sensory stimuli (see dropping arrows!) - and from the skin these converge towards the vertebral column. Running alongside the outgoing motor fibres, but backwards, the lemnisci (sensory bundles) enter as part of the spine’s medulla the foramen, go to the thalamus and then spread into the CNS, its cortical areas.
“Cognition work” begins within the network between incoming sensory patterns feeding into association centers, then converging at the (pre-) motor centres to be released as the outgoing motor patterns.
The senses feed and canalize/constrain behaviour, but some of the sensory patterns “metamorphose” into motor ones – a sensorimotor short circuit, (for) rash action, happening in “mirror”- cells... |
The
spreading and convergence are aspects which in a diagram can only
be shown if it were four-dimensional: the self-channeling function
can be drawn schematically, Fig. 11.
The fountain-metaphor is poor because some important senses are more
linear than surface-like (e.g. the labyrinth), and nothing is explained
on how motor patterns can be constrained by counter-flowing sensory
ones. Actually, there is an area where interaction takes place: the
reticulo-thalamo-cortical system. Multiple interactions by "circuits
within the circuit" are probable, see Fig. 12, see also Hernegger
(1984), who gives a thorough discussion of development of the formatio
reticularis during evolution. |
|
Fig.
11. A self-channeling system.
Fig. 12. Multiple interactions of "circuits within a circuit".
|
|
Another
possibility is a mapping of the dynamical aspects only, the patterns
flowing in various parts of the circuits:
B4. Pattern matching by synchronicity
Motor fibres and sensory fibres conduct patterns in contrary spatial
directions (see above), but can be drawn in parallel as a function
of time. This can be seen from an example how excitation patterns
traveling in a functionally closed system can "match",
which means one part of the circuit is "illustrating",
"commenting" the other ("selfreference").
Imagine
a swimming fish. Because the labyrinth is stimulated only (but immediately)
by acceleration, and the so-called lateral line system in fish monitors
velocity of passing water, we can draw an episode (fish swimming,
increasing and decreasing its velocity, finally hitting an obstacle)
by just parallelling their patterns with the (loco)motor one.
The figure 13 shows (apart from the terrible simplification) how
different senses tell the same story differently. Take for example
the lateral line system, it tells us that after hitting the obstacle
there is still water flowing past the fish. Does that mean the obstacle
is pushed ahead by the fish, thereby slowing him down, or does it
mean that obstacle and fish are situated in a river (the fish perhaps
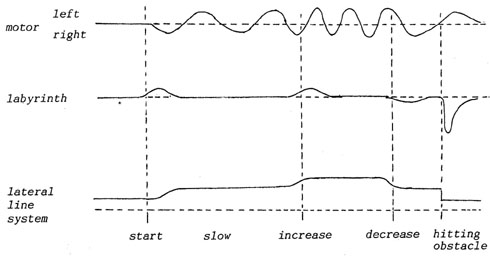
Fig.
13. Correspondence between traveling excitation patterns.
|
|
trying
to smell at the obstacle, continuing to swim to prevent being driven
off?)
One could "ask" the visual and the olfactory sense, and
so can the fish.
Two more things can be shown even in that sketch: Motor patterns
are not altered much by sensory input (say environment), but patterns
of the two senses are strongly dependent on motor acts and environmental
features together! We might assume that dependence of the first
sort will increase by locomotion on a formed and firm substrate
(instead of through amorphous water), which is what J.Gray (1950)
has described.
But how about the second dependence, the inseparability of self-stimulative
and environmental effects in sensory patterns? Will the visual sense,
always regarded as the most accurate in "depicting" the
environment, in "mirroring" it in a "photographic"
way, show that dependence too?
Let
us sketch another episode, but reduce anthropomorphism. First note
that visual afference is dominating amongst our senses. Let us introduce
some crude simplifications:
1. Consider only half-spherical eyes, placed in an exactly lateral
position and built like an insect's eye, Fig. 14
|
 |
Fig.
14. Half-spherical eye.
(from above) |
2.
Assume that there is no eye-muscle activity.
3.
Only two rows of retinal elements are recorded, a horizontal one and
a vertical one, Figure 15. |
 |
Fig.
15. Vertical and horizontal visual detection.
(from the side) |
| 4. Consider
the following situation: Our fish swims slowly and passes both a nearby
and a far-off stationary small object. Simultaneously a swarm of fish
swims against its own direction and one of the fishes passes exactly
the horizontal line of "retinal" elements. Finally our
fish also bumps into an obstacle. |
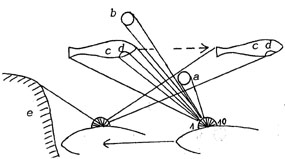
Fig. 16. View from above.
|
a,
b: small objects
c
: other fish
d : eye of fish
e : obstacle
|
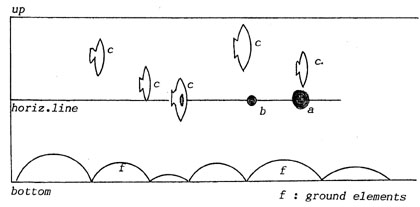
Fig. 17.
"Slot eye", vertical row. |
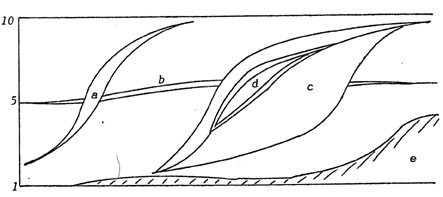
Fig. 18.
"Slot eye", horizontal row. |
|
Apparently
we can identify much easier with the vertical row, probably because
locomotion is perpendicular to it. Anyhow, the sketches show what
complexity the visual system must integrate to represent the environment
as a (pseudostatic) picture.
Two more remarks: Surely eye movements will be used to "follow"
a passing object by stretching it to "real" dimension.
If the fish detects a "homogenous pattern which increases in
a symmetrical way in the frontal part of both eyes", this means
impending collision.
In general, as J.Gibson (1979) has shown, the full two-dimensional
retinal image-pattern changes with locomotion in a regular way:
parts of it leave an one side, other parts enter at the opposite
border, as the "frame" of the retinal image scans along
the environment. So the motor pattern of locomotion is not primarily
represented within the image, but the shift-pattern of the whole
image is "in tune" with it. The pattern of appearance
and disappearance of visual pattern elements does represent time-dependent
properties of locomotion (eye movements excluded).
We are thus led to the conclusion that information about the environment
enters the organism by interfering with an already patterned sensorimotor
"carrier system". Stimuli in this way do not trigger reactive
behaviour, but they are modulating, superimposing on a continuously
interacting organism-environment-system (note 5). External observers
will have access to these processes only when recording techniques
during free locomotioe behaviour are developed.
So far the sensorimotor system was introduced with regard to animals
moving "freely" like a fish. Every muscle action (except
digestion and respiration of course) will influence the relationship
of the whole organism to its environment. But evolution did not
stop with fish.
Part C. Tetrapodes or how to move a marionette
What the selective pressures were that led vertebrates to develop
a neck and legs that carry a body, is still subject of discussion.
Surely orientation and locomotion became accordingly more complex.
In the course of evolution from fish via amphibia and reptiles to
mammals the head became mechanically separated from the rest of
the body and four separated lever systems came to carry an (almost)
passive trunk. New dimensions of behaviour were added by the interposition
of atlanto-occipital, cervical, shoulder, elbow and ankle joints.
The sensorimotor system had to integrate these new internal sources
of disturbance. Let us try to point out a few of the problems to
be solved by an ordered interplay between the anatomical-mechanical
and sensorimotor-behavioural systems. We take mammals as representative:
C1. Posture control
Most of the reptiles pull and push their belly across the substratum.
Mammals have to balance their body, fighting gravity even when standing.
During locomotion certain patterns of gait (walk, trot, canter,
gallop) produce the result of a smooth environmental pass-by (Gibson
1979).
C2. Non-locomotion
Distinctions must be made between motor acts resulting in body or
head movements feeding back environment shift, and non-locomotive
ones like scratching or tail-wagging. Because such non-locomotive
movements are not monitored by vision or touch, another feedback
is necessary:
C3. Proprioception
Proprioception offered the possibility for an internal information
loop. Sense organs within muscles and joints, or next to them give
the CNS "insight" into muscle contractions and joint positions.
Posture and locomotion are now controlled along two pathways: the
orientational circuit and the internal proprioceptive one (note
6).
C4. The reafference principle
Turning the head produces a striking subjective change of the visual
environment (much the same as eye movements) without affecting the
locomotor apparatus. Still orientation is not disturbed! Erich von
Holst and Horst Mittelstaedt (1950) introduced a solution for these
achievements not to be explained by reflex terminology. If there
is no reaction to an environment shift effect following a motor
command, it must be counteracted, extinguished - why not by a "copy"
of the very motor command? Except for a few studies an the "overload-protective"
function of such attenuation of self-induced afferences (Stein,
1978), the importance of the reafference principle might not yet
be adequately understood.
C5. Manipulation
Interestingly, manipulation started with the snout! The principle
is as follows: A paired system of levers is arranged for a coordinated
snapping movement with continuous variability in distance and compression.
Capable of "instant adaption" this pair of pliers has
evolved in fish and has changed anatomical parts since, but gripping,
holding, carrying are functions fully developed mostly in mammals
and birds.
Held objects are integrated in the mechanical body system, they
are "limbs" but without muscular or nervous connectivity.
This means that the existing sensorimotor programs must adapt to
additional loads - new coordinations between labyrinth, neck muscles
and locomotor apparatus have to be, and are established (Horn 1983).
Evolution did not stop with tetrapodes. On the contrary, two ways
of increasing neurocybernetic complexity were successfully followed.
Birds by flight, and primates by tree climbing, really conquered
another, the third spatial dimension. Leaving birds aside we look
for the major consequences of "quadrumanuality" to arrive
finally at the latest condition: bipedal bimanuality.
Part D. Bipedality
Gravity may be the most important single factor in the physical
domain determining anatomical features. For supporting body weight
3 legs would do, but transport without jumps requires 4 ore more.
In case we accept a brachiatory phase in the course of hominization,
"support" of body weight can be achieved by hanging on
one arm, locomotion by augmentation of arms and/or legs (4 of them
plus a tail in some species can be used). Again we can only hint
at some problems for the sensorimotor system:
D1. Tension reversal
We can easily see how mechanical connections remain the same (contact
between limbs and environment) but functional chains ("causality")
change directions. Muscles (in hanging flexors "carry"
body weight, in standing extensors support it) and the sensorimotor
system must rearrange forces and impulses, the labyrinth is forced
to accomodate new, rotatory accelerations.
D2. Spatial abilities and selfmonitoring
Many authors have referred to the importance of eye-hand coordination
in tree climbing and the development of binocular vision for depth
perception, here we need only stress the tight interaction of the
two achievements and their underlying gain in neurocybernetic complexity:
For the first time in evolution animals see what they are doing.
(Canids, carnivores and rodents either lack specialised hands or
cannot afford reducing attention to further environment).
Directly controlled reaching establishes a small, but radically
new range of activity around the animal: Intention movements themselves
can be observed, for the first time the aim and its approach-activities
are monitored, even their contact is observable. Surely direct calibration
of distance is qualitative progress in environment-adapted behavior.
D3. Bipedality
Presently the big apes are by far the most skilled in "instrumental
technology" (ladder building, lever use, sponge use, washing,
throwing, beating, noise production), but of their predecessors
probably only few left the woods and adopted bipedality for locomotion.
The reasons are not yet fully understood, but an interaction of
several factors is probably (Lovejoy 1981). Anyhow, these forelimbs,
- a hundred of million years serving as stabilizing and propelling
fins with "environmental" control, then
- even longer serving as propping legs with additional proprioceptive
control, further
- maybe a dozen million of years serving as suspending arms with
additional visual control were for the first time freed
from stabilizing and Iocomotive functions.
What to do with a pair of legs without regular, firm contact with
the substratum? Not for the first time in evolution did the liberation
of forelegs have positive consequences for cognitive abilities of
the species involved: Arthropods developed antennae for touching
and exploring tasks similar to the use of forelimbs by hominids.
The truly manipulative function, however, is in arthropods performed
by another pair of lifted legs called mandibles....
D4. Hands: pacemakers for hominid cognition?
It is assumed here that arms and hands gained new functions, that
their sensorimotor apparatus, "running free", chose new
interaction partners. Because head and vision left proximity to
the ground, parts of the environment had to be lifted to the manipulative
distance.
Voluntary motor acts had to concentrate an the handled object and
move it with little effort (compared to levering the whole body).
These motor acts which are "individual-bound", "environment-absolute",
established a sensorimotor circuit separated from the locomotion-driven
one. One can say that for forelimbs the environment has shrunk to
a passively moving object, that "behaviour within" had
become "behavior on".
Let us stop here. Bipedality anteceded hominid brain enlargement
as is shown by fossil evidence. Consequences emerging out of the
progressive sensorimotor complication contributed to that development.
Notes
(1) The most common metaphor for spatially complementary structures
in everyday life is the "key-lock" system. One must not
forget that this system works by external forces only: Human cognition
does the selection between patterns, and human hands provide energy
for the mechanical interaction. Yet the spatial logic involved,
the "convexity-concavity-interplay" is comparable on both
macroscopic and molecular level. (At the time when the self-assembling
activity of biomolecules was not recognised, metaphysical "explanations"
in vitalistic terms were given ...)
(2) Euglena may serve as a minimum model for an orientation-locomotion-system.
This famous unicellular organism is propelling itself by flagella
and choosing bright-lightened environments with the aid of a microscopic
eye-spot.
(3) The term Umwelt was coined by von Uexkuell to cover only such
aspects of the environment which are relevant to a single species.
I use environment in that sense, automatically indicating that our
human concepts of environment can differ from the one represented
in various animal species.
(4) The most important feedback of locomotor activity might be the
labyrinth registering change of momentum in any direction. Methodological
problems - the hidden location in the head (any electrodes would
interfere with free locomotion) and the "a priori-like"
information-processing make that sensory mode a remote, "underlying",
inconspicuous one. These characteristics may yet indicate a fundamental
role within the "sensory chorus".
(5) Humberto Maturana in his terminology might say stimuli do trigger
structural changes in the system, but nevertheless conservation
of organization is not endangered by such "perturbations".
(6) Proprioceptive feedback can in a certain way replace the orientational
circuit, e.g. complex nocturnal activity becomes possible. Reptiles
can not use this behavioral niche, but homoiothermal mammals can
(and possibly outdated the dinosaurs in that way).
References
Ashby,W.R. (1952), Design for a brain, Chapman and Hall, London.
Chapeville,F. & Haeenni,A.L. (Eds) (1980), Chemical Recognition
in Biology, Springer, Berlin-Heidelberg, New York.
Gibson,J., (1979), The Ecological Approach to Visual Perception,
Houghton Mifflin Company, Boston.
Gray,J., (1950), The Role of Peripheral Sense Organs during Locomotion
in the Vertebrates, Sym. Soc. Exp.Biol. 4.
Hernegger,R. (1984), Vom Reflex zur Selbststeuerung, Profil Verlag,
München.
von Holst,E. (1969 or 1970 ?), Zur Verhaltensphysiologie bei Tieren
und Menschen I & II, R.Piper und Co, München.
von Holst,E. & Mittelstaedt,H. (1950), Das Reafferenzprinzip,
Die Naturwissenschaften 37, 464-476, 1950.
Horn,J. (Ed.) Multimodal Convergences in Sensory Systems, Gustav
Fischer Verlag, Stuttgart.
Ingenieros,J. (1922), Prinzipien der biologischen Psychologie, Felix
Meiner, Leipzig.
Lorenz,K., (1973), Die Rückseite des Spiegels, R. Piper und
Co, München.
Lovejoy,C.O.(1981), The origin of man, Science Vol. 211, Nr.4480,
341-350.
Maturana,H.(1986?),The Biological Foundation of Self Consciousness
and the Physical Domain of Existence (manuscript).
Roberts,T., (1978), Neurophysiology of Postural Mechanisms, Butterworths,
London Boston.
Stein,P.S.G. (1978), Motor Systems with specific References to the
Control of Locomotion, Ann. Rev. Neurosci. 1, 61-81.
von Uexkuell,J. (1930), Die Lebenslehre, Mueller & Kiepenheuer,
Potsdam.
von Weizsaecker,V.(1940), Der Gestaltkreis. Georg Thieme Verlag,
Leipzig.
Manuscript received: 20-3-1985, final revised version 28-11-1986.
Address of author:
Dr. A. Fuerlinger, Isbarygasse 13, 1140 Wien /Vienna Austria
|
|
|