|
Jeff
Bernard, Gloria Withalm (Hrsg.) Andere
sind das Ziel Einleitung Einleitung
Für
Evolutionsgläubige ist die Frage nach der Menschwerdung aus
dem Tierreich eine der faszinierendsten. Evolutionsbiologie, Verhaltensforschung
und die Neurowissenschaften suchen nach Erklärungsmechanismen,
haben sich aber noch nicht auf gemeinsame Antworten geeinigt.
In diesem offenen Forschungsfeld macht die vorliegende Arbeit
den Versuch, die Steuerung von Wirbeltierverhalten ent-anthropomorphisiert,
möglichst ohne unnötige Vermenschlichung, zu beschreiben.
Ein Beispiel: "Handlungen" sind erst mit den Affen
entstanden, von den Fischen bis zu den Vierbeinern wurde mit dem
Maul manipuliert.
Das
Welterleben von Tieren ohne große Gehirnrinde ist uns unzugänglich,
vermutlich ist es qualitativ anders als unseres. Dennoch lassen
sich für die Bereiche vor der menschlichen Kognition (zu
denen auch unsere vorgeburtliche Entwicklung zählt), Gesetzmäßigkeiten
der Verhaltenssteuerung aus den neuromechanischen Zwängen
der jeweiligen Situation rekonstruieren.
1
Natürliche Kategorisierung
Zur
(Stammes-)Geschichte objektgebundenen Verhaltens
Die
lebenswichtigen Objekte der Tiere sind nicht nur inhomogen in
ihrer Umwelt verteilt, "patchy", sondern sind fast immer
andere Organismen, meist bewegliche andere Tiere (hinfort "Andere").
Da bei uns Menschen anscheinend die Suche nach unbelebten Ressourcen
im Vordergrund steht (Geld, Wohnung, Auto etc. ), müßte
zuerst geklärt werden, ob das Nervensystem (NS), das seit
Jahrhundertmillionen alle Umweltbeziehungen hergestellt hat, nicht
in der Interaktion mit unbelebten Umweltelementen seine
Hauptfunktionen erworben hat.
Methodisch
als eine Art Frage-Antwort-Spiel konzipiert, soll nun von unseren
Objekten in der Gegenwart ausgehend bis ins Paläozoikum zurückgeschritten
werden, um zu den "Urobjekten" bzw. ihren Eigenschaften
zu gelangen.
—
1. Aktuell-situativ gefragt: "Was ist ein Objekt für
Dich, hier und jetzt?" Aufzählung
aller möglichen Dinge, eventuell: "Gibt es ein Mittelding?"
—
2. Individualgeschichtlich gefragt: "Was ist ein Objekt jemals
gewesen - für Dich: Erstes, häufigstes,
wichtigstes?"
Vielleicht
halten sich im "Hier und Jetzt" Personen und Dinge
die Waage, geht man aber im eigenen Leben
zurück, könnten in der Kindheit Geschwister, Kuschel-
oder Haustiere das Auto, den Kugelschreiber oder das Weinglas
an Bedeutung überflügeln. Anschließend
kann
—
3. ethnologisch, anthropologisch gefragt werden: "Was sind
Objekte jemals gewesen - für andere
Menschen, Bekannte, Europäer, Artgenossen; erstes, häufigstes,
wichtigstes?" Die erste
—
4. historische Fragestellung lautet dann: "Was sind bzw.
waren Objekte für die - Großeltern,
Urahnen, "Ötzis"?"
Im ethnologischen Feld sowie beim geschichtlichen Zurückschreiten könnten sich Schmuck, Haustiere, Früchte und Waffen sowie Feinde als wichtig herausstellen. Aber der Großteil der Geschichte fehlt noch, wenn wir "Evolutionäre Psychologie" betreiben wollen. Fragen wir uns —
5. "prähistorisch", nach - Eiszeitmenschen, frühen
Hominiden - und
—
6. "stammesgeschichtlich": "Was machen - Primaten...
Säugetiere... Reptilien... Fische...
erste Tiere"?
Für
die Zeiträume vor der präziser dokumentierten Geschichte
muß Spekulation unser Wissen ergänzen: Tragen frühe
Hominiden neben ihren Kindern vielleicht noch Stöcke und
Steine zum Nahrungszerkleinern, so gehen z.B im Tier-Mensch-Übergangsfeld
Waffen und Instrumente verloren, Affen hantieren fast nur mehr
mit ihresgleichen, Eßbarem, dem Boden und Bäumen.
Vierbeinige
Säugetiere manipulieren fast nur mehr mit dem Maul, tragen
nur Beute und Junge. Ihre Objektkategorisierung nach Wichtigkeit
könnte Feinde, Beutetiere (auch Blätter, Früchte
etc.), Junge, Partner, andere Artgenossen, Wasser, Höhlen
und alle anderen Tiere umfassen, der Rest sind primär neutrale
Biosphären-Elemente, nützlich oder hinderlich erst bei
der Fortbewegung.
Bei
noch früheren, "niederen" Wirbeltieren, etwa
Haiartigen, wird es noch einseitiger, denn alle ihre Objekte
sind Andere oder Teile von solchen: Feinde, Nahrung, Partner,
Konkurrenten. Was sich nicht bewegt, wird oft nicht bemerkt.
Pflanzenfresser-Arten
und/oder -Stadien in der Stammesgeschichte ändern nichts
am Zwang für jedes einzelne Individuum, immer wieder bewegliche
Ziele im Auge zu behalten. Zumindest der Partner, Rivalen und
die Feinde waren selbstbeweglich (und ohne erstere die Stammesgeschichte
zu Ende).
Fazit:
1.
Neurophysiologie ist als "Soziophysiologie" entstanden
(Ein
NS ist an und für sich kaum zu erklären, auch kaum zu
verstehen)
Das
NS ist nach wie vor ein Bewegungssystem, es erzeugt Bewegung und
es reagiert vor allem auf Bewegung, weniger auf (statische) Form.
In der Stammesentwicklung hat die ununterbrochene Wechselwirkung
mit der nicht verhaltensbewegten Umwelt die Leistung der NS ungleich
weniger gefordert, gefördert und geformt als die vergleichsweise
kurzzeitigen interaktiven Verhaltensweisen. Das "Hauptinteresse"
von NS galt daher lange nicht einer stabilen, bewegungslosen Erde
(siehe Kap. 12). Die frühen NS mußten primär ihre
Bewegungen und ihre Beziehung zueinander koordinieren, diese auf
Kosten einer ruhenden Umwelt stabilisieren.
Bei
interaktivem Verhalten treten die beteiligten NS über die
Wahrnehmung von Bewegungseffekten direkt miteinander in Kontakt
und bilden ein sensomotorisch gekreuztes System: Das Verhalten
(die Motorik) des einen steuert über die Sensorik des Anderen
die Verhaltensmotorik des Anderen - und umgekehrt.
Für
ein NS wird also der adäquate Reiz von einem anderen NS erzeugt,
ein NS braucht sozusagen ein anderes, um seine eigenen, wichtigen,
"richtigen", "höheren" Funktionen zu
realisieren und auszuleben. (Katz´und Maus, Katze und Hund,
Katze mit Kater).
Oder,
nach R. Ashby (1970): "Every good regulator of a system
must be a model of that system":
Das
oberste Wahrnehmungs- und Verhaltenssteuerungszentrum, der Homunkulus
der Kognition ist das Verhalten der/des anderen. Andere sind die
ersten "Trainer" oder die (stammesgeschichtlich) angeborene
Lehrmeister - bis der Neocortex diese Rollen übernehmen konnte.
So
betrachtet ändert sich die Darwinsche Einteilung in natürliche
Selektion und geschlechtliche Zuchtwahl zu Kategorien der Nervenfunktionen:
Beute/Räuber-Interaktion, Artgenossen- und Partnerinteraktionen.
2.
Tiere haben kein (leeres) Raumkonzept
Tiere
kennen kein Medium wie Wasser, Luft oder Licht, keine freie Distanz
und daher keinen (Zwischen-)Raum. Ob sie ein von sich getrenntes
Objekt konstituieren können, ist somit fraglich. Eher ist
anzunehmen, daß die Verhaltenssteuerung von Tieren als durchgängige
Sensomotorik, also ohne Ich- und Identifikationsgrenzen abläuft
(siehe Kap. 3). Ihre Fernsinne müßten dann wie Nahsinne
arbeiten, z.B. würde das Verhalten eines Anderen nicht in
unserem Sinne gesehen, sondern im Auge "kitzeln",
für uns eigentlich unvorstellbar.
3.
Unser alltäglicher und wissenschaftlicher Objektbegriff
Die
ursprünglichen Kategorien Beute, Feind, Partner und Neutrale(s)
beherrschen zwar noch immer unsere Emotionalität, das Alltagsobjekt
schlechthin ist aber einfachheitshalber meist starr und unbewegt
(Tisch). Es fehlt ihm die ursprünglich objektkonstituierende
Qualität, nämlich Bewegung. Unsere Nahrung bewegt
sich nicht und Raubfeinde sind entfernt, zwei der wichtigsten
Reizgruppen für Spontanverhalten wirken damit im modernen
Menschen nicht mehr. Pseudobelebte Objekte faszinieren uns (Fußball,
PC), und oft haben wir es mit Zeichen für Bewegung zu tun,
z.B. mit Schrift als Bewegungsspur (siehe Kap. 8), in unserer
Umgebung überwiegen aber leblose Artefakte.
Die
Bühne unseres Einzellebens ist also vollgestopft mit Kulissen,
Versatzstücken und Requisiten. Die Situation ändert
sich erst, wenn Andere auftreten: der Chef betritt das Büro
oder eine Fliege summt in unserer Nähe. Es tut sich eine
Bühne in der Bühne auf, denn Andere verzaubern die Szene
- bis sie identifiziert sind.
2
Die erste Sekunde
Sensomotorische
Kognition in lebenswichtiger Situation ("Aktugenese")
Unter
der Annahme, daß die ersten Fische unserer Abstammungslinie
räuberisch gelebt haben, dabei sicherlich Konkurrenz aus der eigenen und von anderen Arten, vielleicht auch schon Feinddruck ausgesetzt waren, daneben aber auch sexueller
Konkurrenz und Selektion,
hat
sich reflexschnelles Reagieren für alle diese vitalen
Situationen bewährt, auf Verhaltens-"Anzeichen"
hin.
Bevor
es zum Beute- oder zum Sexualkontakt kommt bzw. bevor es überhaupt
zur Jagd oder zur Partnerwahl kommt, muß der mögliche
Beziehungspartner auf Entfernung entdeckt und die Beziehung
hergestellt werden, da Andere oft ihre Präsentation (oder
auch nur Ortung) vermeiden.
Ein
allererster Mechanismus filtert aus dem vielfältigen,
multisensorisch-(eigen-)motorischen "Lärm" einen Geruch, ein Geräusch oder eine Bewegung mit "Hinweischarakter auf relevante Andere" und richtet den Fokus der visuellen Aufmerksamkeit, Augen, Kopf und Körper in Richtung der Quelle. Die Richtung wird mit Peilung festgestellt, z.B. bringt erst die symmetrische Erregung beider Ohren das "Ziel" ins Blick-Suchfeld. Analog dem bekannten Cocktailphänomen filtert emotionales Gestimmtsein bzw. Appetenz das Gewünschte aus dem viel reichhaltigeren Angebot, indem es Eigenaktivitäten hemmt und sensorische Schwellen ganz spezifisch modifiziert. Daten
zur Aktugenese sind rar. Unter Experimentalbedingungen sind die
in freier Natur weitverteilten Reizquellen schwer simulierbar
und im freien Feld scheint eine "kognitive Unschärferelation"
zu gelten: ein Ethologe wird leicht selbst zum relevanten Anderen.
Die
nächstfolgenden Mechanismen müssen sowohl qualitativ
als auch quantitativ verhaltensentscheidend wirken. Auch wenn
schon der allererste geruchliche oder vokal-akustische Eindruck
Aufschluß über die Gruppen-, ja individuelle Identität
des Verursachers gebracht hat, müssen als nächstes Lokalisierung
(Richtung plus Distanz), Bewegungstyp und Größe
der "Ur-Sache" erfaßt werden.
Andere
sind Objekt und Ereignis zugleich. Auge und Ohr können
mit ihrem Bewegungsmuster-Erkennungsvermögen Weg(distanz)
und Ziel(qualität) nebeneinander und gleichzeitig perzipieren.
Die primären Dichotomien lauten "Feind: ja/nein"
und "Beute: ja/nein". Sie stellen die Motorik auf
"vorwärts", "rückwärts"
oder "Abwarten" ein. Emotionen sind dabei der Servomechanismus,
der die bereitstehende Stoffwechselenergie auf breiter Front,
in Millisekunden in Einsatz bringt oder hemmt.
Bei
Tieren entscheiden die dynamischen Merkmale, globale Bewegungsqualitäten
wie Beschleunigungsart und -richtung. Beutetypische Bewegung genügt,
oder feindtypische, um das interaktive Wechselspiel ohne Überlegung
in Gang zu bringen.
Menschen
brauchen zur Kategorisierung und Identifizierung von Objekten
eher statische Eigenschaften, deren Wahrnehmung und Vereinigung
zu einem Begriff mindestens eine halbe Sekunde kostet. Bei Tieren
sind in dieser Zeit viele Kämpfe ums Dasein schon dem Ergebnis
nach entschieden, viele auch schon beendet.
Die
Entdeckungssituation erfaßt aber (in Millisekunden !) nicht
nur den gesamten eigenen sensomotorisch-emotionalen Apparat,
die eigene Reaktion kann zur ebenso raschen gegenseitigen
Identifikation führen. Aktion, Re-aktion, Re-re-aktion werden
dann so eng ineinandergreifen, daß vom ersten Blick an beide
Seiten in einem System gefangen sind: Motorik des Einen
stimuliert die Sensorik des Anderen, mobilisiert seine Motorik,
diese stimuliert wieder die Sensorik des Einen, usw.
Dieser
Metakreislauf der Reiz-Reaktions-Folgen ist nicht wie ein Dialog,
als Kommunikation, als Frage-Antwort-Spiel zu beschreiben. Für
Codierungen sind die Zeiträume zu kurz und in der Interaktion
überschneiden sich die Intervalle. Es ist ein reflexschneller
Wettlauf der Sensomotoriken auf Leben und Tod. (Bei der Partnerwahl
findet ein - eher langsamerer - Tanz statt, aber mit ebenso
exakter Mikrosynchronisierung der komplementären Bewegungsmelodien).
"Emergency
reaction" heißt wörtlich überrasches Auftauchen
und unmittelbarer Reaktionszwang. Die erste Phase der Interaktion
von Sensomotoriken ist der Höchstleistungsmechanismus
nervöser Funktion und als solcher einer der Kernmechanismen
natürlicher Selektion überhaupt. Es wäre auch schwer
einzusehen, daß sich für jeden der anfangs aufgezählten
Kontexte ein eigener neuraler Mechanismus (Modul) entwickelt.
Bei
der Jagd werden Individuen zwar gejagt, aber nicht erkannt.
Individuelle
Identifikation findet nur im innerartlichen Bereich statt,
als Partnerwahl und männliche Konkurrenz im Kontext sexueller
Selektion und ab den Säugetieren (und Vögeln) als Lehr-
und Lernverhältnis in der eusozialen Brutpflege. (Im Verhältnis
zu den Jagd- und Fluchtkontexten vergleichsweise in Zeitlupe).
Die
ersten "vollständig erkannten Individuen" in
der Stammesgeschichte könnten übrigens Männchen
gewesen sein, da sie sowohl von Weibchen als auch von konkurrierenden
Geschlechtsgenossen unter Fitnessaspekten, d.h. auch "ganzkörperlich-dynamisch"
betrachtet werden. Für die karge Ressource, die Weibchen
für Männchen darstellen, genügen attraktive Einzelmerkmale.
3
Über die Grenzen individuellen Verhaltens
Fragmente
einer erweiterten Nerven-System-Theorie
1.
Ein Gehirn ohne periphere Nerven ist ein Artefakt
2.
Ein Nervensystem ohne Muskeln ist unfähig zur Bewegung
3.
Ein Nerv-Muskel-Präparat kann ohne kohärente Körpermechanik
nur zucken
4.
Ein Tier ohne Umweltwiderständigkeit bleibt immobil
5.
Ein mobiles Tier hat ohne Andere niemals ein Ziel.
ad
1: Sensomotorik
Wie
das Herz ist das Gehirn auffälligster Teil eines Kreislaufsystems.
Harvey hat gezeigt, daß die Riesenmenge Blut, die das Herz
passiert, nicht im Körper verbraucht und neu gebildet werden
kann. Auch im Nervensystem werden die Riesenmengen an motorischen
Impulsen und sensorischen Erregungen nicht unabhängig voneinander
produziert und verbraucht, bei Tieren in Bewegung werden sensorische
Erregungen primär von der eigenen Motorik verursacht.
Die
individuelle Sensomotorik arbeitet als funktioneller Kreislauf,
das durchgehende Prinzip sind Wirk-Muster. ("Muster stehen
zwischen Chaos und Ordnung", C. Bresch 1977).
Wirkmuster
sind die Übertragungseinheiten, sie durchwandern und wechseln
dabei ihre Trägerstrukturen, dafür behalten sie aber
ihren zeitlich-räumlichen Wirkzusammenhang bei. Die dynamischen
Charakteristika der Muster, z.B. Rhythmus und Melodie, schließen
die Umwelt in den Funktionskreis ein: Motorikmuster »
Bewegungsmuster (entsprechend Beziehungsänderungen
seitens der Körperteile bzw. Umweltteile) » Sensorenzustandsänderung
» Sensorikmuster » ZNS »
Motorik usw.
ad
2: Ohne Muskeln gäbe es kein Nervensystem
Da
die ersten Muskelzellen noch selber erregbar waren, müßten
auch alle späteren Sensorensysteme letztlich dem (An-)Trieb
dienen. Muskeln setzen die linear-exekutiven Erregungsmuster in
analoge Zug- und Druckeffekte um, die Wirkmuster divergieren
im Körper. Zum Teil betreiben sie die Festkörpermechanik
des Skelettsystems als Stützen, Hebel, Zange etc., zum Teil
münden sie in die nichtfeste "wetware", die ihren
eigenen, hier selten gewürdigten Gesetzlichkeiten folgt (siehe
nächster Punkt).
ad
3: Konstruktions-Morphologie
Ein
Individuum ist unteilbar in dem Sinne, daß Eröffnung
seiner Hohlräume den Tod bewirken kann. Sein kohärenter
Körperverbund gehorcht, von der einzelnen Zelle bis zum reißfesten
Hautsack der Wirbeltiere, dem hydraulischen Prinzip,
das den mechanischen Versteifungen sowohl unter- als
auch übergeordnet ist (W. Gutmann 1988). Peristaltik, Atmung,
Greifbewegungen, eine Umarmung können nicht linear-kausal
beschrieben werden, erst die unhierarchische, aus allen Richtungen
konvergierend gleichen Gegendruck vermittelnde Hydraulik,
die ineinander eingesackten Kompartimente geben dem üblichen
Marionettenmodell der Wirbeltiere Ganzheitlichkeit, Masse und
gefüllte Form. Tiere sind nicht flächig modellierbar.
ad
4: "Muskel bewegt die Welt" (R. Granit 1977)
Innerhalb
der Biosphäre hat jede Körperoberfläche eine Grenzfläche
zu den Umgebungsmedien wie Wasser, Luft und/oder Boden. Sich bewegende
Körper werden Wechselwirkungen hevorrufen, geringe mit der
Luft, aber auch unnachgiebige mit dem Erdboden. Eine wichtige
Wechselwirkung für Tiere bringt ihre Lokomotion hervor: Die
Lokomotorik bewegt das gesamte Gewicht des Körpers. Aber
wenn sich ein Körper bewegt, bewegt sich relativ zu ihm auch
seine Umgebung: Für die tierische "Lokosensorik"
bewegt sich die ganze Welt.
ad
5: Für Tiere grenzt das eigene Verhalten an das der Anderen
Wenn
zwei Tiere sich bewegen und ihre Umwelten dabei verschieben, dann
wird aus der Erde und den zwei mobilen Tieren auf ihr ein Dreikörpersystem,
in dem es keinen Punkt in Ruhe gibt. Die visuelle Welt zwischen
Tieren gehört funktionell für sie zum je eigenen Körperbewegungssystem.
Andere sind von vornherein ein Element der lokosensorisch bewegten
Welt, ruhend können sie als Teil dieser (Um)Welt unsichtbar
bleiben. Erst ihre Eigenbewegung wird auf sie aufmerksam machen
und sofort zum kontrastreichsten und informativsten Merkmal werden
(s. Kap. 2).
Das
heißt: Der Umriß des Anderen ist jeweils die Grenze
der unterschiedlichen Bewegungseffekte, nicht die eigene Haut.
Das
heißt weiterhin, daß Tiere zwischen sich und Anderen
kein Konzept einer ruhenden Welt haben oder brauchen. Für
einen menschlichen Beobachter, egal ob er auf der Erde ruht oder
sich auf ihr bewegt, ist ebendiese Erde stabil und in Ruhe. Dazu
einige allgemeinere Betrachtungen.
4
Exkurs über Stabilität
Zur
Entstehung von Fläche, Linie und Punkt
Eine
Fläche entsteht zwischen zwei qualitativ verschiedenen,
einander begrenzenden Räumen.
Ein
Beispiel: die Erdoberfläche. Sie wird sehr kompliziert, wenn
man sie genau nimmt und Strukturen wie Bäume samt Blättern
einbezieht.
In
der Biosphäre ist jede Oberfläche eine Grenzfläche.
Sie kann eine Trennfläche sein, ein Interface z.B. zwischen
Innen und Außen. Sie entsteht auch oft zwischen hydraulischen
oder pneumatischen Kompartimenten, durch gegenseitigen Druck.
Als Modell können zwei Seifenblasen dienen, die aneinander
"kleben".
Eine
Linie entsteht durch konvergenten Druck dreier (annähernd
symmetrischer) Kompartimente.
Ein
Beispiel: die Küstenlinie. Sie bildet sich zwischen Wasser
und Festland und der Luft darüber.
Im
Modell: drei Seifenblasen können sich nur in einer Linie
treffen.
Bewegung,
z.B eine einfache Translationsbewegung, erfordert Kanalisierung.
Die lineare oder Richtungs-Determination geschieht durch
Druck von außen, die Bewegungs-Linie ist das Produkt mindestens
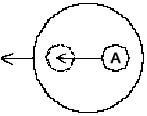
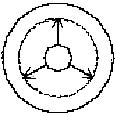
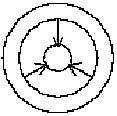
dreifach-seitlicher Einschränkung. Ein Pfeil in Bewegung
braucht, wie auch manche Haie zeigen, mindestens drei seitliche
"constraints" (Abb. 1, 2).
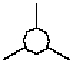 
Abb. 1
Abb. 2
Um
einen (Körperschwer-)Punkt im Raum in Ruhe, stabil
zu halten, müssen seine Bewegungsmöglichkeiten allseits
eingeschränkt werden. Die für uns denkübliche "Einschachtelung",
also Druck aus sechs Richtungen ist möglich, aber mechanisch
überdeterminiert.
Um
ein makroskopisches Objekt im Raum zu fixieren, braucht Druck
nur aus vier Richtungen gleichzeitig und konvergierend
ausgeübt zu werden. Um das "unkooperativste" Objekt,
eine harte glatte Kugel, festzuhalten, reichen zwei gekreuzte
Zangen mit tetraederartiger Druck"flächen"verteilung
bzw. -konvergenz aus. Der "Konvergenz-Tetraeder" ist
das einfachste Modell für die Stabilisierung und damit auch
Manipulation eines Objekts. Er stellt auch das einfachste Hydraulikmodell
dar. Der (Ruhe-)Punkt ist vierfach constrained, ein höchstdefinierter
Ort.
Will
man auf der Erdoberfläche einen Körper ruhen lassen,
braucht er drei Standpunkte. Damit sind schon drei mögliche
Bewegungsrichtungen "constrained", die nach unten,
eine vierte (die nach oben) wird durch das Gewicht, durch den
Zug der Schwerkraft, verhindert. Dreibeinige Tische, viele Pflanzen
und (meeres)bodennahe einfache Tiere zeigen diese Gesetzlichkeit
in ihrer dreifach-radiären Symmetrie. Mehr zu dieser "Raumlogik",
einer Spielart der Geometrie, bei A. Fürlinger (1987 u. 1989).
5
Verhalten als Drei-Körper-Problem
In
Lokomotion ist der Weg eines Individuums Resultante zweier Attraktoren,
eines physikalischen (Gravitation zur Erde) und eines biosozialen
(Attraktion, Repulsion durch Andere). Bei einem landlebenden Individuum
stehen die Attraktionsziele, die Richtungen zu ihnen, grob rechtwinkelig
zueinander: Erde unten, Andere vorne. Das Dreikörperproblem
stellt sich von außen, für einen Beobachter als Vierten,
ganz anders dar als für ein beteiligtes Individuum von innen.
von außen: 1,2: Individuen von innen:
3: Erde
(cf. besser: Abb. 10)
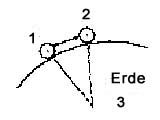
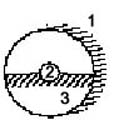
Abb. 3 Abb.
4
Das
Bauprinzip, mit dem mechanisch, sensorisch und motorisch die Richtungsstrahlen
zur Erde und zu Anderen innerhalb des Individuums gemeinsam kontrolliert,
geführt werden können, ist die Bilateralsymmetrie: Weg
und Ziel liegen in einer Ebene, der virtuellen Ebene zwischen
links und rechts.
Darüberhinaus,
d.h. auf die Umwelt bezogen, hat jedes Verhaltenssystem seinen
eigenen Richtungs-Konstanz-Mechanismus. Schwerkraft und soziale
Interaktion arbeiten gleichartig, indem seitliche Abweichungen
durch Körperbewegungen innerhalb eines engen Sensorenbereichs
gehalten werden. Zuerst ein fester Stand, dann der Blick auf andere
(cf. Kap. 6).
Die
Beziehung zur Erde ist für einen Landbewohner dann
ruhestabilisiert, wenn die Lotrechte (die Verbindungslinie der
Körperschwerpunkte beider Körper) innerhalb ihrer Kontaktflächen
gehalten wird. Jeder Schwerpunkt stellt die Spitze einer Stabilitätspyramide
dar, "auf" einer gemeinsamen Kontaktfläche, d.h.
auf minimal drei (Eck-) Punkten:
1 Individuum
3 Erde
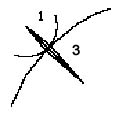
Abb.
5
Dies
gilt auch für alle Körper, die auf Stelzen ruhen,
z.B. (vom Inneren der Erde aus gesehen):
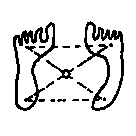  
Abb. 6
Abb. 7
Abb. 8
2-Beiner 1-Beiner Zehensteher Während
der Lokomotion kann sich die Lotrechte kurzzeitig außerhalb,
meist vor der Kontaktfläche befinden; um einen Sturz zu vermeiden,
muß sie aber "eingefangen" werden, beim Stehenbleiben
der Menschen zB. durch Zurücklehnen oder einen längeren
Schritt.
Der
Zwang zur Geschwindigkeitserhöhung z.B. bei der Jagd führt
durch "run-away selection" für schwimmende Körper
zur Stromlinienform, bei landlebenden zur Verkleinerung der Kontaktflächen
und Verminderung ihrer Anzahl.
Die
Extremform der Fortbewegung zu Lande wäre die lineare Aneinanderreihung
von Kontaktpunkten wie beim Radfahren und Seiltanzen. Der Stabilitätstetraeder
bildet sich zwischen einer Unterstützungskante, die längs
verläuft, und einer queren Oberkante, die als Lenkstange
oder Balancierstange das Gewicht ausbalanciert.
Für
die Lokomotionsorgane bedeutet all dies extreme Selektion auf
Symmetrie, denn sie verbrauchen viel Energie, und Fehler addieren
sich schnell zu "deviantem" Verhalten. In der sexuellen
Selektion dient daher gute Symmetrie ("Schönheit")
als wichtiges Kriterium, es wurde für Insektenflügel
und die Beine von Rennpferden nachgewiesen.
6
Blickverhalten
Analog
zur Lotrechten, die über die Somästhetik der Beinmuskulatur
und der Fußsohlen die Haltungs-, Stellungs- und Lokomotionsmotorik
steuert, ist die visuelle Orientierung zu Anderen ein Management
der virtuellen "Beziehungslinie" zwischen beiden Körpern.
Da
es den Blick-Abstand als leere Distanz für Tiere nicht gibt,
wird aus der relativen Größe des Anderen zusammen mit
den jeweilig sichtbaren Bodenelementen bis zum Ziel eine motorische
Weg-Distanz geschätzt werden, wie dies J. Gibson (1982)
beschreibt. Die Ausrichtung und die Korrekturen der Blickrichtung
beruhen auf Körperbewegungen, mindestens der Augen und/oder
des Kopfes.
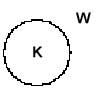
In
subjektiver Sicht schneiden sich die Symmetrieebene (der Weg)
und die Horizontlinie (der Erdrand) im Ursprung (origo) unseres
cartesianischen, rechtwinkeligen Koordinatensystems :
Rand des Blickfeldes = Körper (1) Horizontlinie (3) Objekt/Anderer (A)
Bodenelemente, Weg (W)
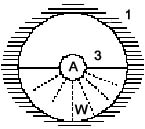
Abb. 9
Wenn
sich das Objekt/Andere bewegt, wird das Auge seitlich oder sagittal
"nachgeführt":
Ernst
Mach war vielleicht der erste, der in der Skizze "Das visuelle
Ich" diese Situation veranschaulichte, allerdings mit dem
eigenen Körper im Zentrum seiner Umwelt:
 Abb.
10
Der
Blick auf den eigenen Körper ist allerdings eine stammesgeschichtlich
späte Situation, entstanden durch zwei Krümmungsbewegungen
der Körperachse im Laufe der Wirbeltierevolution: Bei den
Fischen sind Lokomotion (der Vortrieb durch die Schwanzflosse)
und visuelle Zielorientierung noch in einer Linie angeordnet.
Bei der Landnahme durch die Vierbeiner schwenkt der Lokomotionsapparat
um 90 Grad nach unten, Labyrinth und Auge müssen den Körper
über den neuen flexiblen Hals fernsteuern (Vögel auch).
Affen schließlich beugen die Körperachse im Halsbereich
weiter, beim Sitzen vertikal nach unten. Die Extremitäten
reichen nun nach vorne und gelangen damit zum ersten Mal auf Dauer
ins eigene Blickfeld (siehe Abb. 10 und Kap. 12).
Bevor
aber der eigene Körper ins Bild kommt, soll er den Rahmen
für seine Umwelt abgeben.
7
Exkurs: "Endo-neurologie"
Zur
Geschichte der Darstellung von Weltbildapparaten
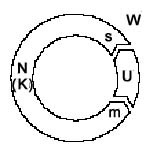
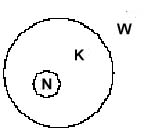
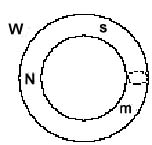
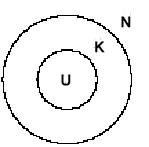
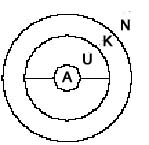
Legende:
W = Welt, ext. Beobachter
Eine Darstellung der subjektiven Sicht braucht eine relativistische Umstellung der Beteiligten (analog zur Beschreibung der "Endophysik" von O. Roessler 1992). Das ergibt dann:
Begründung
des Endo-neuro-Modells:
Das
wahrnehmende und handelnde Subjekt (be)findet sich objektiv und
subjektiv innerhalb des Bereichs des Nervensystems. Seine Sensoren
und motorischen Synapsen umgeben es, als Interface zwischen Innen-
und Außenwelt. Bei Säugetieren z.B. sind die taktilen
Rezeptoren fast körperbedeckend angeordnet, sie repräsentieren
das Interface als ausgedehnt und flächig geschlossen, wie
einen Käfig. Am dichtesten sind sie in den Sinnesorganen
angeordnet, als die Fenster, durch die sich das Individuum am
meisten Umweltreizung beschafft.
Trotzdem
umgibt der nichtneuronale Körper das Nervensystem vollständig.
Die äußere Hautschicht vermittelt letztlich, als "weiteres"
Interface, zwischen den Welten (beim Menschen zwischen mentalem
Raum und Umwelt). Der Körper stellt sich als Ring oder Torus
dar, er könnte gleichzeitig als geschlossen und offen, als
konvex und konkav betrachtet werden (ob ein Interface konvex oder
konkav ist, kann von einer Seite her nicht bestimmt werden).
Die
Abb. 6 bis 8 sowie 16 zeigen nun, wie Lotrechte
und Blick, die zwei verschieden gelagerten Teilprobleme der Verhaltenssteuerung,
mit geringem sensomotorischen Aufwand richtungsstabil gehalten,
d.h. in der Zeitachse kanalisiert werden können. In beiden
Mechanismen umfassen, manipulieren Körper(teil)bewegungen
die Beziehungsachsen.
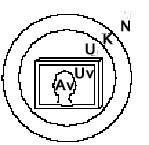
Die
Situation eines PC-Users bei der Arbeit stellt sich übrigens
noch komplizierter dar:
8
Lokomotion macht Geschichten
(Fortbewegung
als Vor-gang, Pro-cedere, Pro-zeß, Fort-Schritt)
Tiere
haben mit der Fortbewegung die ewige Suche gewählt. Unter
allen Verhaltensweisen ist die Hin- und Fortbewegung, die gerichtete
Lokomotion, die älteste und energieaufwendigste. Ihre Steuerung
von der Zielvorgabe bis zum mechanischen Ablauf mußte zu
allen Zeiten, in allen Individuen in unendlich variierenden Situationen
erfolgreich verlaufen (Be-Weg-ung).
Die
Summe aller Orte, die ein Individuum nacheinander aufsucht, ergibt
seinen je eigenen Lebensweg. Nimmt man den Körperschwerpunkt
als virtuellen Repräsentanten des ganzen Individuums, kann
man die Bewegung dieses Punktes, die Kurve die er in Raum und
Zeit zurücklegt, als Lebenslinie bezeichnen. Sie beschreibt
von oben gesehen die aktuelle geographische Bewegungsrichtung
des Individuums, seitlich gesehen zeichnet sie die Bodenunebenheiten,
das Profil des Weges, nach (beim Menschen etwa in Bauchhöhe).
Im
Sinne des Nerv-Muskel-Körper-Umwelt-Zusammenhanges produziert
die Lokomotion nicht nur eine virtuelle Linie, sie reiht gleichzeitig
und lückenlos die einzelnen Verhaltensepisoden hintereinander.
Lokomotorik und -sensorik selektieren so den Weg als Folge
von Situationen im Sekundenbereich aus der Umwelt, für das
Individuum, aber auch für die Art und den (Wirbeltier-)Stamm.
So
kann man sagen, auf dem Weg von den Fischen bis zum Menschen hätten
die einzelnen Situationen als Episoden praktisch alle paar Sekunden
gewechselt. Es hat sich deshalb bewährt, drei Zeitskalen
zu konstruieren, von denen jede in der nächsten als Mikroelement
enthalten ist:
1. Aktugenese, Situation, Ereignisgeschichte (cf. Kap. 2) 2. Individualgeschichte, Ontogenese (s. Lebenslinie) 3.
Stammesgeschichte (Phylogenese) s. Abb. 18
Übrigens
hat Höherentwicklung bei den Wirbeltieren bis zum Menschen
wirklich stattgefunden. Fische steuerten durchs dreidimensionale
Meer, Amphibien stiegen ans Land, Säugetiere hoben
den Körper auf Stelzenbeine; Affen hoben die Vorderbeine,
zogen den Vorderkörper in die Höhe, kletterten
auf und in die Bäume, Hominiden hoben den Vorderkörper
samt Armen permanent in die Höhe. Menschen schließlich
schießen einander auf den Mond:
 Abb.
18
Lokomotion
zieht sich als "Roter Faden" durch alle Geschichten,
nicht nur durch die biologischen. Sofern sich eine Geschichte
durch Raum und Zeit bewegt ("storyline"), wird sie
explizit oder implizit nach dem elementarsten Bewegungsprogramm
beschrieben, das wir haben.
Wenn
es ein autobiographisches, ein "narratives" Gedächtnis
gibt, läuft es über den Lokomotionsapparat, der funktionell
nicht vom übrigen Körper getrennt werden kann.
Die
"Reise des Helden" begann mit den ersten Tieren und
deren Suche nach Nahrung, Schutz und Partnern. Sie endet, "vor-läufig",
mit der Literatur, wo die Schreibenden mit ihren Vorderbeinen
immer noch kleine motorische Spuren ziehen ...
Die
älteste und wichtigste Bewegungsform der Wirbeltiere muß
auch die Grundlagen für ihre Orientierungsparameter bieten.
Erst die Körperschwerpunktsteuerung kann der Lokomotion "Sinn"
geben, eine Koordination der Körper-Teilbewegungen "im
Sinne" der stabilen Führung herbeiführen. Die
resultierende Lebenslinie kann dann vom Auge und den mechanischen
Sinnen kontinuierlich und synchron "mitgeschrieben"
werden, als illustrierte Bewegungsmelodie. Den "Grund"-Rhythmus
geben dabei die lokomotorischen Muskelaktionen mit wechselnden
taktilen Hautsensationen.
Vestibularsystem
und Taktilität werden ununterbrochen, "millisekündlich
durch die Jahrzehnte" über den Bewegungszustand von
Körper und (ge)wichtigen Körperteilen samt Erdwiderstand
informiert und schalten sich niemals ab.
Die
Hypothese dazu lautet: Solange das gesamte Körpergewicht
dynamisch prozessiert wird (Lokomotion, aktive Haltung, Stellung),
untersteht das ganzkörperliche senso-motorische Orientierungssystem
dem Labyrinth, auch das Auge ge"horcht" dem Innenohr
(E. v. Cyon 1908).
Sind
Raum und Zeit bei Tier und Mensch gleich?
Tierische
und spontan unbewußt menschliche Sensomotorik entsteht aus
Bewegung, perzipiert Bewegung und reagiert darauf mit Bewegung.
"Blindsehen" bei Affen und Menschen zeigt, daß
auch ohne primäres Sehzentrum adäquat reagiert werden
kann.
Woher
kommen Raum und Zeit, wenn Bewegung zur Verhaltenssteuerung ausreicht?
K. Lorenz sagte einmal sinngemäß, daß Raum und
Zeit Unterkategorien von Bewegung seien. Das hieße, erst
der Mensch hat sie durch Zerlegung von Bewegung gewonnen, sie
sind nicht Grundkategorien, sondern unsere Beschreibungsweise
der Urgegebenheit Bewegung.
9
Die flüchtige Gestalt
Im
Anschluß an die "erste Sekunde" (s. o.) nun
zum faszinierendsten und vielleicht wichtigsten Verhalten unserer
Vorläufer, der Jagd.
Wenn
in der Entwicklung zum Menschen die Beziehung zwischen Einzelindividuen
oft in Jagen und Gejagtwerden bestand, ist auch unserem Nervensystem
diese unbeschreibliche "mechanisch-kognitiv- emotionale
Drei-Körper-Problem-Situation" funktionell tiefer verankert
als jede andere.
Ob
unsere Fischahnen jeweils größere Raubfeinde um sich
hatten, ist nicht sicher, daß sie aber selber Räuber
waren, schon. Für die ersten kleinen Säugetiere können
wir jedenfalls in den Dinosauriern wichtige Verfolger vermuten
(Drachenmythos), sicher wichtigere, als es Säbelzahntiger
und Höhlenbären in der Menschwerdungsphase waren.
Die
Verhaltenssteuerung bei der Jagd ist für Jäger und Gejagte
derart komplex, daß die sensomotorischen Systeme zur Reduktion
ihres Agierens auf Relevantes, also zu Konstanzmechanismen gezwungen
werden: Die Lokomotionsbewegungen werden auf Automatik gestellt,
d.h. Geländedetails nur im Notfall visuell beachtet. Im Blickfeld
wird auch nur das Wichtigste stabilisiert. Für beide Teilnehmer
ist das weder das detailgenaue Äußere des Anderen,
auch nicht dessen (Körperteil-)Bewegungen (!), sondern die
gezielte Manipulation der Beziehung zu ihm, d.h. das Erreichen
bzw. Aus-den-Augen-"Verlieren" des Anderen. In extremer
Bewegung, während sich Richtung und Profil des Weges permanent
ändern, kann vom Anderen nur die mittlere anatomische Grundstruktur,
die "flüchtige Gestalt", verfolgt werden.
Der
eindeutig zentrierbare/permanent bewegte Rand des Anderen
ist "die Gestalt" des Anderen.
Es
soll hier nahegelegt werden, daß das Komplexitätsreduktions-
Prinzip, das während voller Eigenbewegung die "Identität"
des Anderen garantiert und seinen "visuellen Schwerpunkt"
im Auge behält, unserem Gestaltprinzip in der Psychologie
funktionell zugrundeliegt. Die Interaktion kann damit unter wechselnden
Sicht-, Licht- und Projektionsbedingungen weiterlaufen, auch wenn
der Andere nur unvollständig oder als (Teil)Umriß im
Blickfeld bleibt.
Die
Gestalt entspräche damit einem "permanenten Bestätigungsmechanismus
im (dyadisch-)interaktiven Blickkontakt während großer
sensomotorischer Beanspruchung".
10
Verfolgen: Die erste mimetische Fertigkeit
Die
Neigung unseres NS, extern gesteuert zu werden, ist um vieles
älter als die, Andere zu manipulieren. Der Begriff "Machiavellian
Intelligence" (Byrne/Whiten 1988) bezeichnet nur letztere
und läßt den "passiven" evolutionären
Vorlauf der Jagdkontexte außer Acht.
Schon
die frühesten Jäger unserer Stammesgeschichte koppelten
ihre Sensorik an die Motorik der Beute. So konnten sie "on-line"
jede kleinste Kursänderung des Verfolgten zur Organisation
der eigenen Motorik einsetzen und waren damit erfolgreich:
 Abb.
19 (nach Driza/Cholewa 1985)
Das
Nachmachen der Lebenslinie eines Tieres im sensomotorischen Verbund
durch Andere soll "Mimesis Typ I" genannt werden. Eine
Ähnlichkeit der annähernd parallelen Trajektorien ist
nur für Dritte, externe Beobachter wahrnehmbar (Abb. 19).
Ein Jäger macht ja nicht die Einzelbewegungen des
Beutetieres nach - er hat ja meist eine andere Anatomie, sicher
auch eine andere Motivation - , er koordiniert seine Bewegungen
aber dergestalt, daß er wie durch ein unsichtbares
Band von seiner Beute (an)gezogen erscheint. Die Beute wiederum
muß ihre Konstanzmechanismen unter gemischt-entgegengesetzten
Vorzeichen steuern, denn die Geländebedingungen kommen von
vorne und der Verfolger von hinten.
Ein
anderes Beispiel: Durch die "Blume" wird dem jungen
Hasen gesagt, wo es lang geht. Der Schwanz des Muttertiers ist
sein Führungselement, sein locus of control. Es gibt Tempo
und Richtung vor, aber das Nachfolgen geschieht aus eigenem Erkennen
und aus eigener Kraft. K. Lorenz hat auch Gänsekinder "angeführt",
die auf seine Gestalt geprägt waren. Für Beobachter
sieht es wie Kind-er-ziehen aus, aber ohne Zug und ohne
Druck.
Das
Fangen-Spielen jugendlicher Säugetiere zeigt vielleicht
am besten die Untrennbarkeit von Sensorik, Motorik, Individuen,
Phylo-, Onto- und Aktugenese: Die schnellen Folgen von Perspektiven-und
Rollenwechseln, und bei Gefahr auch Strategiewechseln, erlauben
es jedem Individuum, die phylogenetisch perfektionierten mimetischen
Fertigkeiten in aktuellen (Lern-)Situationen einzuüben.
Bei
der innerartlichen Konkurrenz (Definition der Konkurrenz:
mehrere laufen zusammen auf ein Ziel) um Territorien, Nahrung,
Sexualpartner, aber auch während Balz und Brutpflege, stehen
baugleiche, oft verwandte Tiere in permanenter Wechselsteuerung.
Für
die Semiotik ist mimetisches Verhalten, wie es bisher beschrieben
wurde, nicht leicht zu definieren, da erst mindestens zwei Individuen
zusammen das Phänomen hervorbringen. Die Vorgangsähnlichkeit
könnte man "kinetisch-ikonisch" nennen, um der
fast synchronisierten Doppelbewegung als einer Art Resonanzphänomen
gerecht zu werden.
Was
bei Tieren im sensomotorischen Verbund abläuft und im Schwarm
hunderte Individuen synchronisieren kann, ist beim Menschen vor
allem in zwei Bereichen noch wirksam: beim Tanz und im militärischen
Drill (McNeill 1995). Außerdem kann man bei Boxkämpfen
das Phänomen der Ideomotorik beobachten, man sieht die Zuschauer
mit ihren Körpern "mitgehen".
Imitation
und Pantomime finden normalerweise nicht im sensomotorischen Verbund
mit dem Modellverhalten statt und sind Fähigkeiten des Nachmachens
aus dem Gedächtnis. Bevor wir solche Produkte "fertigen"
Menschseins betrachten, gilt es, den Übergang von den Vierbeinern
zu den Vierhändern zu beschreiben, die Affenwerdung.
11
Der Griff
Als
Schrittmacher für den Lauf der Wirbeltierevolution waren
die Extremitäten immer gewohnt, den ganzen Körper zu
bewegen, sei es als steuernde, rudernde Flossen oder als schiebende,
laufende, kletternde Beine.
Sie
"veränderten" ihre Umwelt durch die Aufeinanderfolge
aufgesuchter Nischen.
Für
Fische war und ist die Erde äußerst nachgiebig und
homogen, für Landbewohner ist sie teils nicht vorhanden (Luft),
teils relativ(istisch) unbeweglich, stabil (Erdoberfläche).
Für
Primaten schließlich ist der Untergrund schwankend geworden:
Bäume sind zwar formfest, aber ihre Äste werden mit
Distanz vom Boden zunehmend schmal und biegsam.
Da
Äste für die Beine von Tieren "teilweise verdeckte,
glatt-elastische, in alle Raumrichtungen geneigte und gekrümmte
Unterstützungslinien zwischen lauter Abgründen"
darstellen, wurde zu ihrem Besteigen ein neuer Kontaktmodus hervorgebracht,
der Griff.
Seit
Fischzeiten hat das Maul der Wirbeltiere seine Koppelungs- und
Fixationsaufgaben erfolgreich gelöst. Nur in seltenen Fällen
demonstrieren Vogelschnäbel die (instabile) Zwei-Punkt-Zange.
Bei anderen Tiergruppen drücken Zähne, Zunge, Gaumen
und Lippen in komplexen Raummustern zu. Die "Brechzange"
der hundeartigen Tiere zeigt übrigens den Stabilitätstetraeder
für harte Objekte: vier Druckkanten, je zwei an einem Zahn
des Ober- und Unterkiefers, einander überkreuzend.
Hände
entwickeln darüber hinaus qualitativ und quantitativ
neue Eigenschaften:
—
Exploration: Sie sind Sinnesorgane, gleichzeitig fern- und kontaktgesteuerte
Sonden, "Meßfühler".
—
Delegation, relative Autonomie: Ihr Wirksystem arbeitet dem Körper
gegenüber (1) in variabler Distanz,
(2) cardanisch, d.h. fast in jeder Lage, und
(3) mit lockerer oder stangenartig steifer Verbindung.
—
Kooperation: Mit Maul und Schwanz können bis zu sechs Koppelungen
im Raum installiert werden.
—
Manipulation: Erst zwei Stabilitäts-Systeme können ein
Objekt frei bewegen, z.B. mehr als 360
Grad rotieren.
Als
Koppelungsmodus arbeitet der Griff "schließlich"
mehr als mechanisch, nämlich als "leere" oder
"offene Hydraulik". Greifen ist stufenlos regelbar
in
— Richtung, Geschwindigkeit und Dauer, — Spannweite ("Umarmung" bis zum Kapillarspalt zwischen zwei Finger- kuppen), — Umschließungsgrad (Haken bis Öse, Mulde bis geschlossene Höhle), — Reibung (Hautfeuchtigkeit), und 12
Alles ruht?
Die
Sensomotorik für Baumbewohner wie Primaten spielt sich überwiegend
nah und frontal ab, weil wichtige Umweltstrukturen in allen anderen
Raumrichtungen "wegfallen":
—
Äste bieten Lokomotionsangebote, die nach "vorne"
divergieren oder zur Umkehr zwingen,
—
Blätter bzw. Nachtaktivität schränken den Sehraum
ein,
—
Bäume sind funktionelle Sackgassen für größere
Raubtiere.
Das
"Gesicht", aber vielleicht noch mehr die aufgerichtete
Körperhaltung machen Primaten uns ähnlich. Durch überwiegende
Gewichtsverlagerung zwischen die Hinterbeine und Sitzen scheinen
schon die Affen die berühmte Aufrichtung grund-"setzlich"
vollzogen zu haben.
Beim
Sitzen ist die Labyrinthfunktion zwar nicht "entspannt"
wie beim großflächig-stabilen Liegen, aber doch von
leitenden und tragenden Koordinationen entlastet, eben "sediert"
(=beruhigt).
Die
Folgen des Lokomotionsverzichts sind sensationell: Die Welt
steht still. Ruhe.
Eine
gewagte Hypothese: Der Eindruck einer ruhigen, feststehenden Außenwelt,
"die Situation", wie wir Menschen sie erleben, begann
mit dem Sitzenbleiben der Affen.
Begründung:
Das bis zu den Primaten einheitlich arbeitende System zur Körperbewegung
(Blicklinie in Verlängerung der Körperachse, siehe Kap.
6) kann sich im Sitzen funktionell aufspalten: Hinterbeine und
das Hinterteil bilden eine Unterstützungsfläche, auf
der der Körper mit der Erde verschmilzt. Das Labyrinth, das
bisher den Körper indirekt manipuliert (besser pedipuliert)
hat, braucht erstens nur mehr den Kopf, also sich selber zu balancieren.
Es kann zweitens den ganzen Körper samt Schwerpunkt fast
in einer Linie unter sich kontrollieren. Diese Anordnung entläßt
die Vorderbeine und Augen aus dem Körpermanagement.
Erst
die Entlastung der Sensomotorik beider Systeme, Augen und
Vorderbeine, kann zwischen diesen ein relativ autonomes, rekursiv
geschlossenes Nervensubsystem hervorbringen. Der "Auge-Hand-
Komplex" schafft qualitativ neue (vom Boden abgehobene, freie,
absolute) sensomotorische "Eigen"-schaften, die auch
beim Aufrechtgehen erhalten bleiben. Sind sie der Beginn eines
"Identität"-Erlebens im Sinne von Selbsterkenntnis
des eigenen Körpers? Das Subjekt muß doch zuerst Objekt
werden, um unter Objekten zum Besonderen (= eigenen) zu werden.
Hände sind Objekt und Subjekt(-teile) zugleich.
Mit
den Hominiden ist für die Savanne eine Verhaltens-Chimäre
entstanden: Die obere Körperhälfte ist damit beschäftigt,
die (Dinge aus der) Welt zwischen Augen und Händen auf neue
Art zu begreifen. Die untere will mit dem alten, nie vergessenen
Laufprogramm wieder Anschluß an flüchtige Bodenbewohner
gewinnen...
13
Mimesis Typ II
Die
Suche, Entdeckung und Einverleibung von Eßbarem war bis
zu den Primaten "reine Kopfsache", Nase, Ohr und Auge
haben das Maul zum Ziel gesteuert. Der Körper fungierte dabei
als gleichsam ausgelagerter, ferngesteuerter Servomechanismus.
Die
Verfolgung von Kleinlebewesen mit Arm und Hand ist sensomotorisch
und emotional eine völlig neue Art von Jagd, ein Neu-an-"Fang",
das Modellverhalten für "Mimesis Typ II":
—
Das Gesamtkörper-Nachfolgeprogramm (Mimesis Typ I) wird innerhalb
des Körpers delegiert, die
Orientierungs-Kontrolle durch den Kopf bleibt (ruhig), aber
das Zuschnappen ist Sache der Hand.
—
Meist macht nur eine Hand den An-griff, Händigkeit
ist eine Folge der Jagd, bei der normalerweise
ein bewegliches Ziel die sensorischen und
motorischen Prozesse kon-zentriert. —
Da der Energieaufwand der Armbewegung im Vergleich zur Ganzkörperjagd
gering ist, ist auch die Motivation geringer:
In Reichweite der Arme kann beliebig gekoppelt
und entkoppelt werden, auch ohne die emotionale
Balance zu verlieren. —
Das Vorausschicken der Arme, ihre Vorläufigkeit, d.h. freie
Reversibilität, in allen Richtungen
läßt echtes Probehandeln zu. Das Testen von
Ästen auf Festigkeit oder das Heranholen
von attraktiven Früchten stehen meist nicht unter
starkem Zeitdruck.
—
Die eigenen Bewegungen erhalten Handlungs-Spiel-Raum, der Zeitrahmen
wird disponibel, ein Bereich der Überlegung,
aber auch der Willkür eröffnet sich.
—
Die Greifdistanz ist für Tiere die erste direkt nachvollziehbare
Raumdistanz, sie kann später
in der Menschwerdung durch Werfen vergrößert werden.
Eine
weitere Hypothese: Die Konzentration auf das Beute-Hand-Geschehen
läßt den Hintergrund, der mit dem Körper ruht,
vergessen. Die Greifbewegungen mögen dann unversehens auf
dem eigenen Körper oder auf dem des Nachbarn landen. Die
manuelle Jagd kann sich so zur sozialen Fellpflege ("grooming")
verbreitern. Im Partner wird durch die zarten Hautreize eine beruhigende,
ruhigstellende Hormonausschüttung ausgelöst.
Soziales
Berühren scheint doch, zumindest für uns Menschen, ambivalent
besetzt: Annäherung zum (Haut-)Kontakt muß erst als
Nichtangriff gekennzeichnet werden (K. Lorenz 1963). Da aber An-greifen
(phylogenetisch erst mit der Primatenhand entstanden) ontogenetisch
mit dem Klammern nach der Geburt beginnt, wird auf taktiler Kontaktbasis
sicher früher Vertrauen gelernt als Mißtrauen (Eibl-Eibesfeldt
1984).
Bei
sozialen Berührungen können weder das Auge noch die
Pfote/Hand allein zwischen den eigenen und fremden Körperteilen
unterscheiden, erst die andere Seite, die Sensibilität des
be-troffenen Hautareals, entscheidet. Ohne frühes Wissen
vom eigenen Körper ist das entscheidende Identitätsgefühl
schwer zu erklären.
Die
Identifikation von "eigen" (im Unterschied zu "fremd")
ist notwendig subjektiv.
14
"Woman is ... the first real mover" (C. Paglia 1990)
Da
der ontogenetischen Frühsituation eine echte Grundlage fehlt
(Säugetiere beginnen ihr Leben schwebend, als schwerelose,
blinde "Amnionauten"), stülpen sich als Ersatz
für den noch nicht vorhandenen Bodenkontakt zwei Hautareale
am embryonalen Kopf nach innen und bauen sich kleine "Erdmodelle"
(Statolithen, allseitig sinneszellenerregend fixiert). Zusammen
mit den cartesianisch-dreidimensional gebauten Bogengängen
liefern sie als Lage- und Kinetikdetektoren die ersten Impulsmuster
an das selbstorganisierende Nervennetz.
Dieses
braucht für das Anlegen seiner späteren Funktionen viele
"Informationen", sprich Abwechslung, und wenn möglich
in sinnvoller, gemusterter Komplexität. Schön wäre
ein Verhaltens-Lehrmeister in utero.
Wir
alle hatten einen solchen, die Alltagsbewegungen des Mutterkörpers:
Sie gehen den eigenen, ebenfalls lageverändernden Muskelzuckungen
des Embryos a priori voran.
Auch
am Beginn des neuronalen Lebens, in der "Ontogenese",
wirkt also eine andere individuelle Motorik als Organisator der
Neurodynamik. Man könnte sie Mimesis Typ III nennen:
Der
Fetus macht als passiv bewegter Körperteil alle Körperbewegungen
der Mutter mit, ihren Gehrhythmus, ihre Art zu tanzen, zu sitzen,
ihre Umdrehungen im Schlaf. Bevor wir denken, haben wir eine mehrmonatige
Reise im physikalischen Schwerpunkt der Mutter mitgemacht und
mitgelebt. Wanderungen, Auto- und Schifahrten, Stiegensteigen,
kaum eine menschliche Bewegungsweise ist uns nicht vertraut, bevor
wir in die Lage kommen, einen Blick auf die Welt zu werfen oder
mühsam den ersten Schritt zu tun.
Ein
vorgestellter Synchronvergleich der (taktil-)labyrinthären
Afferenzen von der Mutter und ihrem Fetus, ihrer kinetischen "Fahrtenschreiber",
ergäbe einander ähnliche, allerdings nicht identische
Musterfolgen. Der körperzentral gelegene kindliche Kopf liefert
im Vergleich zu dem der Mutter nur Rumpfdaten, globale Übereinstimmungen
wären trotz dieses relativen Filtereffekts zu erwarten.
Die
Erfahrenslinie des Labyrinths vereint doch alle relevanten Hebe-,
Senk- und Lateralbeschleunigungen sowie Rotationen des gesamten
Trägersystems. Kann sie auch "ohne Körper"
die für jedes Individuum exklusive, einzigartige Bewegungsspur
in Raum und Zeit, die Lebenslinie, darstellen? Die Kopfposition
einer Person stellt den wandernden Schnittpunkt der physikalischen,
biologischen und soziokulturellen Welt dar, weil sie in jedem
Moment allen "herrschenden" Bedingungen (physikalischen
und biologisch-organischen Gesetzmäßigkeiten, individuellen
psycho-sozialen Einflüssen) gehorcht...
Weiters
wäre anzunehmen, daß kleinere technische Transportmittel
wie Auto, Boot oder Flugzeug vorgeburtliche Bewegungsmuster annähernd
simulieren und primär beruhigend wirken können.
Die nachgeburtlichen visuellen Eindrücke könnten die
labyrinthären Vor-erfahrungen bildhaft illustrieren und damit
bewußtseinsfähig machen, z.B. für unsere archaischen
Schwebeträume.
15
Du hast dich selbst längst erkannt
Da
dem Hautorgan zu Beginn der Ontogenese ein Dialog mit Anderen
verwehrt bleibt (außer bei Mehrlingen), tritt es, sanft
durch die Kuppel der Eihäute unterstützt, mit sich selbst
in Kontakt. Wenn mit der Körperdifferenzierung die sprossenden
Extremitäten lang genug geworden sind, um rekursiv auf den
Körper zurückgebeugt zu werden, und wenn das Nervennetz
hautflächendeckend geworden ist, kommt es zur "Selbst"(=Doppel)-Berührung.
Anders
als das Bewegungsmuster nachvollziehende Labyrinth beginnt die
Hauttaktilität, ein räumliches Weltmodell aufzubauen,
und zwar mit den Objekten die vor-handen sind, mit den eigenen
Körperteilen.
Eine
letzte Hypothese: Die "Homunkuli" in den sensomotorischen
Cortices entstehen ohne Störung von außen, intrauterin,
in den praktisch pausenlosen Kontaktereignissen der Selbstumarmung.
Sie sind am größten, d.h. am detailreichsten dort,
wo anatomiegemäß die häufigsten Kontaktereignisse
statt"finden": zwischen Händen und Gesicht.
Die
ontogenetische Koordination der Greifmotorik mit rein taktiler
Perzeption ist die Innenseite des phylogenetischen Grooming-Programms.
Sie ist erst für Säugetiere notwendig, weil sich erst
bei diesen unsere kleinsten Lehrmeister, parasitische Andere,
im Fell verstecken können, also unsichtbar wurden.
Um
nach der Geburt einen Reizpunkt (Mückenstich) auch blind
zu orten, z.B. durch gezieltes Kratzen, müssen vorher alle
potentiell betroffenen Reizpunkte einzeln ansteuerbar sein. Der
"Zugriffsplan" sind die Homunkuli, die (späteren)
Meßfühler Hände nehmen schon bei den ersten Gelegenheiten
am eigenen Körper das sensomotorische Maß aller (späteren)
Dinge. Auch das Auge wird, etwa in der späteren Greifpraxis,
primär dem "blind"-exakten sensomotorischen Wissen
zu Diensten sein, vor dem projektiv-überlegenden Distanzschätzen.
Die
Unterscheidung zwischen eigenen Körperteilen und Fremdstrukturen
ist qualitativ eindeutig: letztere haben nur ein taktiles
"Echo" (seitens des Fühlers).
Die
innere Eihaut als äußerer Tasthorizont ist zwar in
allen Richtungen vor-handen, sie ist eine Biosphäre von innen,
aber doch ein konkaver, glatter und endloser Widerstand ohne faßbare
Strukturen. Als Trost für die "ungreifbare" Mutter
gibt es ein echtes erstes Objekt der Ontogenie, die Nabelschnur.
Sie hat die richtige Größe und genügend Freiheitsgrade
für Manipulation und gibt auch nur ein taktiles Echo.
Überschreitungen
des Tasthorizonts, "Übertritte", können
nach außen durchschlagen, von anderen gespürt werden.
Sie fordern von Mutter und Dritten die existentiell wirklichsten
persönlichen Dimensionen ein: (an)erkannten und selbst verteidigten
Eigen(spiel)Raum. Vertreibung daraus ist ab diesem Zeitpunkt
Kampf.
Epilog
Eine
Wissenschaft beschreibt die Wege des Lebens, auch den Weg durch
die Jahrhundertmillionen bis zum Menschen: die Biologie.
Viele
Wissenschaften beschreiben den Menschen: Anthropologie, Medizin,
Psychologie, Soziologie, "Hirn"forschung, Linguistik,
Ethnologie, Semiotik. Eine allerletzte Hypothese:
Das
Interface aller Wissenschaften vom Menschen, der gemeinsame Nenner,
ist die Biologie.
Danksagung
Ich
möchte L. Deecke, K. Edlinger, W. Gutmann, D. Ploog, R. Riedl,
G. Schweigart, M. Wimmer und E. Vanecek für kritische Anmerkungen,
A. Schmitt für seine hilfreiche Begleitung und G. Withalm
& J. Bernard für die reichliche Überarbeitungszeit
danken. Dies ist daher auch der Ort festzuhalten, daß es
sich inhaltlich seit ca. 1994 gegenüber dem Vortrag aus 1990
um eine de facto neue Version handelt.
Literatur
Bresch,
C. (1977). Zwischenstufe Leben. Evolution ohne Ziel. München
- Zürich: Piper
Byrne,
R. & Whiten, A. (1988). Machiavellian Intelligence.
Oxford: Clarendon Press
Conant,
R.C. & Ashby, W.R. (1970). "Every good regulator of
a system must be a model of that system". International
Journal of Systems Science 1-2: 89-97
Cyon,
E. v. (1908). Das Ohrlabyrinth als Organ der mathematischen
Sinne für Raum und Zeit. Berlin: Springer
Driza,
M & Cholewa, G. (1985). Leben und Umwelt. Lehrbuch für
Biologie und Umweltkunde. Band 1 für die 1. Klasse AHS, für
die 1. Klasse HS. Salzburg: Verlegergemeinschaft Neues Schulbuch
Eibl-Eibesfeldt,
I. (1984). Die Biologie des menschlichen Verhaltens. München:
Piper
Foerster,
H. v. (1986). Sicht und Einsicht. Braunschweig: Vieweg
Fürlinger,
A. (1987). "Ortungen. Raum — Auge — System".
In: Bernard, J. (Hg.). Semiotica Austriaca. Wien: ÖGS,
56-60
Fürlinger,
A. (1989). "Eins-Zwei-Drei". In: Koch, W.A. (Hg.).
Das Ganze und seine Teile. Bochum: Brockmeyer, 89-96
Gibson,
J.J. (1982). Wahrnehmung und Umwelt. Wien: Urban &
Schwarzenberg
Granit,
R. (1977). The Purposive Brain. Cambridge, MA-London: The
MIT Press
Gutmann,
W.F. (1988). "The hydraulic principle". American
Zoologist 28: 257-266
Lorenz,
K. (1963). Das sogenannte Böse. Wien: Dr. G. Borotha-Schodler
Maturana,
H. (1982). Erkennen: Die Organisation und Verkörperung
von Wirklichkeit. Braunschweig: Vieweg
McNeill,
W. (1995). Keeping together in Time. Cambridge, MA: Harvard
University Press
Paglia,
C. (1990). Sexual Personae. New York: Vintage
Roessler,
O. (1992). Endophysik. Die Welt des inneren Beobachters.
Berlin: Merve Verlag
Uexküll,
J. v. (1928). Theoretische Biologie. Berlin: J. Springer
|